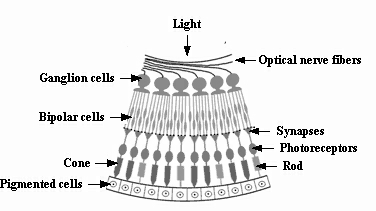
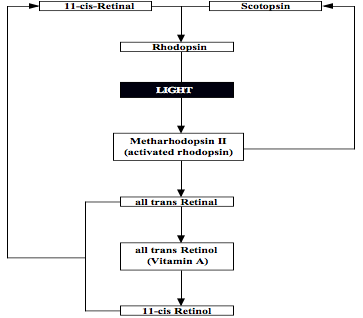
Two classes of photoreceptors in the retina. In total, there are over 125 million of such receptors in the retina, with the rods being more numerous at some 120 million than the cones at around 6 to 7 million (see first figure below). Both of these photosensitive membranes have distinctive shapes, both are stacked in orderly pile of discs littered with pigment transmitted from cells beneath them, and they synapse at their other ends with bipolar (and horizontal) cells in the retina, to which they transmit nerve impulses (see figures below). The cones, responsible for high visual resolution and color perception, are classified into three types of color receptors, each containing different visual pigments, and having broad and overlapping response (or spectral sensitivity) curves ‘tuned’ to different portions of the visible spectrum based on measures of peak absorption: 1. red-absorbing cones (64%) that absorb optimally at relatively long wavelengths peaking at 565 nanometers (nm), 2. green-absorbing cones (32%) with a peak absorption at 535 nm, and 3.blue- absorbing cones (2%) with a peak absorption at 440 nm, which have the highest sensitivity and that are found mostly outside the fovea. Their responses are not all-or-none, as a light of a given color wavelength (e.g., 500 nm for green) stimulates all three types, but in this example the green-absorbing cones will be stimulated most strongly. The rods, responsible for scotopic or dark-adapted vision, monochromatic vision, and motion detection, are more than a thousand times as sensitive as the cones such that they can be triggered by individual photons under optimal conditions. Optimal dark-adapted vision is attained only after 30 minutes or longer of darkness, as the rod adaptation process is much slower than that of the cones. New discs are added to the base of the outer segment of the rod cell at a rate of 1 to 3 per hour, as the old ones are shed off at the tip, and become phatgocytosed by the pigment cells that also contain vitamin A. They contain a photosensitive pigment called rhodopsin or visual purple, which is a mixture of the protein scotopsin and 11-ciss-Retinal that is derived from vitamin A, the latter accounting for why a deficiency in this vitamin can create visual problems (see second figure below). When activated by light, rhodopsin triggers an electrical impulse that travels to the ganglion cells and then to the optic nerve for eventual transport to the primary visual cortex. At present, the cellular pathways and regulatory molecules that determine exactly how photoreceptors develop are poorly understood, especially with regard to rods, perhaps because of their relatively small number. Like all neurons, however, retinal cells must develop according to an interplay of intrinsic and extrinsic cues. Thus, for example, to become a rod, a retinal progenitor cell has to encounter a set of environmental cues, but at a time when it has the ability to respond.