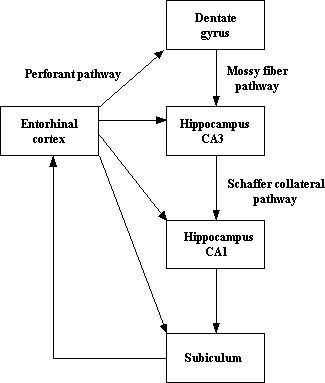
Located in the rostral half of the ventromedial surface of the temporal lobe, the entorhinal cortex (EC) is part of the hippocampal formation, with the other areas being the hippocampus proper, the dentate gyrus and the subiculum. Its name derives from the fact that its location is inside the rhinal sulcus in the olfactory area. Consisting of six layers, although Ramón y Cajal (1832-1952) identified seven layers in the 15-day-old human fetus, it is classified as area 28 according to Brodmann’s system. The EC has widespread output connections with many cortical, subcortical and hippocampal structures (some of the latter being illustrated in the first figure below). Two-thirds of the cortical projections to the EC are by means of the parahippocampal and perihinal cortices, and most are reciprocal, with the former receiving inputs from dorsal stream and the latter from the ventral stream (see second figure below). However, the olfactory structures are the only primary sensory systems that have direct projections with the EC. It also has a number of subcortical connections: cholinergic fibers of the basal forebrain project to the entorhinal cortex, emotionally-controlled information is received, and from the brain stem, serotonergic afferents to the entorhinal cortex originate in the raphe nuclei, and noradrenergic inputs arise from the locus coeruleus and ventral segmental area. Furthermore, thalamic afferents innervate the EC. Being the origin of the major afferent fiber system to the hippocampus via the perforate pathway, and the interface between the hippocampus and the cortex, the EC is depicted as playing a crucial in memory, and in particular declarative or long-term memory. Other functions accorded to the EC include allocentric spatial representations (in the dorsocaudal medial entorhinal cortex) and the regulation of blood glucose. When damaged or diseased, it has been implicated in, for example, temporal lobe epilepsy and Alzheimer’s disease as well as schizophrenia as a consequence of a diminution in the number of its neurons. Whether pathology of the EC plays a role in schizophrenia is more controversial as little is known about dopaminergic innervation of this structure in the primate brain. Unfortunately, little is known also about the normal structural and functional development of this amazing part of the hippocampal system in the human.